Tin chuyên ngành
on Tuesday 21-07-2020 9:17am
Danh mục: Vô sinh & hỗ trợ sinh sản
ThS. Võ Như Thanh Trúc – Chuyên viên phôi học – IVFAS
- Giới thiệu
- Cơ chế phân tử của quá trình trưởng thành nhân noãn
(1) GVBD – germinal vesicle breakdown:
Sự tiếp tục quá trình giảm phân của noãn được khởi phát để đáp ứng với tác dụng của hormone LH (Luteinizing hormone) ở tuổi dậy thì khi noãn bắt đầu tích tụ dịch nang. Noãn lúc này sẽ nhận hormone progesterone từ các tế bào cumulus qua các thụ thể của hormone này gắn trên màng sinh chất của noãn. Khi các hormone đã hoàn toàn tiếp xúc với thụ thể, cytoplasmic domain của thụ thể khởi phát một tín hiệu phân tử nội bào làm hạn chế sự tổng hợp cAMP nội sinh [1]. Ngoài ra, một nghiên cứu đã cho thấy sự tiếp tục quá trình giảm phân của noãn còn được điều hỏa bởi sự ngừng tương tác giữa noãn và các tế bào granulosa [4]. Trong nghiên cứu này, người ta tách noãn ra khỏi các tế bào granulosa và nhận thấy các noãn này thoát khỏi sự bắt giữ chu trình tế bào tại Prophase I và tiếp tục giảm phân. Như chúng ta đã biết, noãn và các tế bào này tương tác với nhau bằng tương tác gap junction (cầu nối liên bào) và các tế bào cumulus liên tục cung cấp cAMP cho noãn. Chính vì thế, khi các tương tác này giảm đi, lượng cAMP cung cấp cho noãn cũng giảm đi đáng kể. Kết quả của cả hai hiện tượng trên làm giảm nồng độ cAMP nội bào. Nồng độ cao cAMP nội bào dẫn đến kích hoạt yếu tố ức chế phiên mã PKAR/c, yếu tố này kiểm soát không cho noãn phiên mã các protein cần thiết để tiếp tục chu trình tế bào, chính vì thế ở giai đoạn GV chu trình tế bào của noãn bị bắt giữ ở prophase I. Khi nồng độ cAMP giảm, yếu tố ức chế phiên mã PKAR/c bị bất hoạt, noãn tiếp tục phiên mã dịch mã các protein đặc trưng, trong đó, 2 protein CyclinA và Cdk2 tham gia hình thành phức hợp protein CyclinB/Cdc2 đồng thời phosphoryl hóa phức hợp này (do bản chất của Cdk2 là một kinase protein). Sau khi được phosphoryl hóa, CyclinB/Cdc2 hoạt động như một kinase protein, làm phá vỡ túi mầm (GVBD).Sở dĩ CyclinB/Cdc2 có thể làm tan túi mầm (màng nhân noãn) là vì màng nhân của noãn cấu tạo chủ yếu bởi các protein lamin kết hợp với nhau (một nghiên cứu sử dụng các kháng thể kháng các lamin này đã xác định được màng nhân noãn được cấu tạo bởi 2 loại protein – lamin B và lamin AC) [3], [6]. Nhưng khi sự phosphoryl hóa quá mức, các lamin rời nhau ra, màng nhân tan biến. Dưới kính hiển vi quang học, ta có thể nhận biết GVBD qua hiện tượng mất màng nhân noãn (hình 2.1).
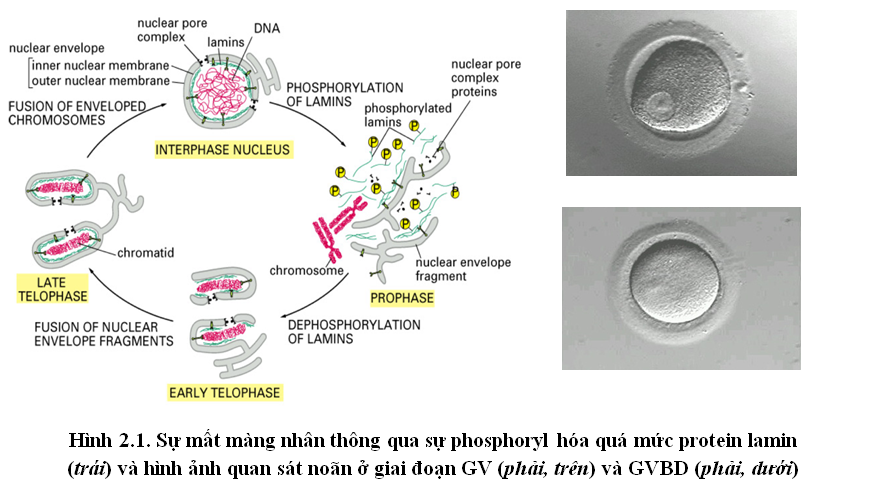
(2) Các NST (nhiễm sắc thể) đóng xoắn:
Một số nghiên cứu cho thấy sự phosphoryl hóa protein histone H3 ở vị trí Ser10 (đuôi amino) tạo thuận lợi cho các yếu tố hỗ trợ đóng xoắn (được gọi là các condensin) tiếp xúc với sợi nhiễm sắc. Condensin là một phức hợp gồm 5 protein được bảo tồn trong quá trình tiến hóa, bao gồm nhân SMC (structure maintenance of chromosomes) ở dạng heterodimer và 3 tiểu phần non-SMC (hình 1.2) [2]. Như vậy, ở cuối prophase I, NST đã đạt trạng thái đóng xoắn cực đại.
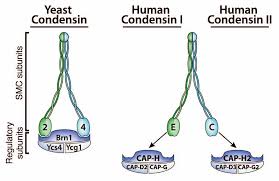
Hình 2.2. Cấu trúc condensin ở người [5].
TÀI LIỆU THAM KHẢO
1. Anja Schmitt, A.R.N., Signalling pathways in oocyte meiotic maturation. Journal of Cell Science, 2002. 115: p. 2457 - 2459.
2. Franck Bazile, J.S.-P., Damien D'Amours, Three-step model for condensin activation during mitotic chromosome condensation. Cell Cycle, 2010. 9(16): p. 3263 - 3275.
3. Maul G., S.G., Jimenez S., et al., Detection of nuclear lamin B epitopes in oocyte nuclei from mice, sea urchins, and clams using a human autoimmune serum. Devel Biol, 1987. 121: p. 368 - 375.
4. Racowsky C., B.K., In vitro and in vivo studies reveal that hamster oocyte meiotic arrest is maintained only transiently by follicular fluid, but persistently by membrana/cumulus granulosa cell contact. Devel Biol, 1989. 134: p. 297 - 306.
5. Seang Lin Tan, R.-C.C., William M Buckett, In-vitro maturation of human oocytes: Basic science to clinical application. 2007.
6. Stuurman N., H.S., Aebi U., Nuclear lamins: their structure, assembly, and interactions. J. Struct Biol, 1998. 122: p. 42 - 66.
Mời xem tiếp phần 2
Các tin khác cùng chuyên mục:
TIN CẬP NHẬT
TIN CHUYÊN NGÀNH
LỊCH HỘI NGHỊ MỚI
Năm 2020
New World Saigon Hotel, thứ bảy ngày 17 tháng 01 năm 2026
Năm 2020
Hội Nội tiết Sinh sản và Vô sinh TP. Hồ Chí Minh (HOSREM) sẽ ...
Năm 2020
Khách sạn Equatorial, chủ nhật ngày 23 . 11 . 2025
GIỚI THIỆU SÁCH MỚI

Ấn phẩm CẬP NHẬT KIẾN THỨC VỀ QUẢN LÝ SỨC KHỎE TUỔI MÃN ...
Y học sinh sản được phát ngày ngày 21 . 9 . 2025 và gởi đến ...
Cẩm nang Hội chứng buồng trứng đa nang được phát hành online ...
Hội viên liên kết Bạch kim 2024





Hội viên liên kết Vàng 2024

Hội viên liên kết Bạc 2024

FACEBOOK